一 : 细胞因子:细胞因子-概述,细胞因子-重组要求及状态
细胞因子(cytokine)是指主要由免疫细胞分泌的、能调节细胞功能的小分子多肽。在免疫应答过程中,细胞因子对于细胞间相互作用、细胞的生长和分化有重要调节作用。由于基因工程、细胞工程研究的飞速发展,不仅克隆了早先发现的生物活性肽的cDNA,而且发现了许多新的细胞因子,并对各种细胞因子产生来源、分了子结构和基因、相应的受体、生物学功能以及与临床的关系等进行了大量的研究,成为当今基础免疫学和临床免疫学研究中一个活跃的领域。
细胞因子_细胞因子 -概述
细胞因子(cytokine)是由机体多种细胞分泌的小分子蛋白质,通过结合细胞表面的相应受体发挥生物学作用。1、细胞因子的命名细胞因子有多种其他的名称:如由单个核吞噬细胞产生的细胞因子称为单核因子(monokine);由淋巴细胞产生的细胞因子称为淋巴因子(lymphokine);可刺激骨髓干细胞或祖细胞分化成熟的细胞因子称为集落刺激因子(colonystimulatingfactor,CSF)等。由不同细胞分泌的细胞因子,其基因及编码蛋白与功能均清楚者,通常均归属于白细胞介素(Interleukin,IL)。白细胞介素、干扰素、肿瘤坏死因子、生长因子和趋化性细胞因子都是细胞因子。在固有性免疫应答及适应性免疫应答过程中,细胞因子发挥重要的功能。细胞因子_细胞因子 -重组要求及状态
重组细胞因子要求:细胞因子_细胞因子 -分类
细胞因子是多种细胞所分泌的能调节细胞生长分化、调节免疫功能、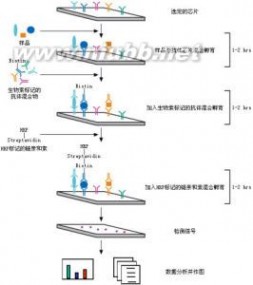 细胞因子(抗体芯片)参与炎症发生和创伤愈合等小分子多肽的统称。免疫球蛋白、补体不包括在细胞因子之列。
细胞因子(抗体芯片)参与炎症发生和创伤愈合等小分子多肽的统称。免疫球蛋白、补体不包括在细胞因子之列。(一)根据产生细胞因子的细胞种类不同分类
1.淋巴因子(lymphokine) 于命名,主要由淋巴细胞产生,包括T淋巴细胞、B淋巴细胞和NK细胞等。重要的淋巴因子有IL-2、IL-3、IL-4、IL-5、IL-6、IL-9、IL-10、IL-12、IL-13、IL-14、IFN-γ、TNF-β、GM-CSF和神经白细胞素等。
2.单核因子(monokine) 主要由单核细胞或巨噬细胞产生,如IL-1、IL-6、IL-8、TNF-α、G-CSF和M-CSF等。
3.非淋巴细胞、非单核-巨噬细胞产生的细胞因子 主要由骨髓和胸腺中的基质细胞、血管内皮细胞、成纤维细胞等细胞产生,如EPO、IL-7、IL-11、SCF、内皮细胞源性IL-8和IFN-β等。
(二)根据细胞因子主要的功能不同分类
1.白细胞介素(interleukinIL) 1979年开始命名。由淋巴细胞、单核细胞或其它非单个核细胞产生的细胞因子,在细胞间相互作用、免疫调节、造血以及炎症过程中起重要调节作用,凡命名的白细胞介素的cDNA基因克隆和表达均已成功,目前已报道IL-1-IL-15。
2.集落刺激因子(colonystimulatingfactor,CSF) 根据不同细胞因子刺激造血干细胞或分化不同阶段的造血细胞在半固体培养基中形成不同的细胞集落,分别命名为G(粒细胞)-CSF、M(巨噬细胞)-CSF、GM(粒细胞、巨噬细胞)-CSF、Multi(多重)-CSF(IL-3)、SCF、EPO等。不同CSF不仅可刺激不同发育阶段的造血干细胞和祖细胞增殖的分化,还可促进成熟细胞的功能。
3.干扰素(interferonIFN) 1957年发现的细胞因子,最初发现某1种病毒感染的细胞能产生1种物质可干扰另1种病毒的感染和复制,因此而得名。根据干扰素产生的来源和结构不同,可分为IFN-α、INN-β和IFN-γ,他们分别由白细胞、成纤维细胞和活化T细胞所产生。各种不同的IFN生物学活性基本相同,具有抗病毒、抗肿瘤和免疫调节等作用。
4.肿瘤环死因子(tumornecrosisfactorTNF) 最初发现这种物质能造成肿瘤组织坏死而得名。根据其产生来源和结构不同,可分为TNF-α和TNF-β2类,前者由单核-巨噬细胞产生,后者由活化T细胞产生,又名淋巴毒素(lymphotoxin,LT)。2类TNF基本的生物学活性相似,除具有杀伤肿瘤细胞外,还有免疫调节、参与发热和炎症的发生。大剂量TNF-α可引起恶液质,因而TNF-α又称恶液质素(cachectin)。
5.转化生长因子-β家族(transforminggrowthfactor-βfamilyTGF-βfamily) 由多种细胞产生,主要包括TGF-β1、TGF-β2、TGF-β3、TGFβ1β2以及骨形成蛋白(BMP)等。
6.趋化因子家族(chemokinefamily) 包括2个亚族:(1)C-X-C/α亚族,主要趋化中性粒细胞,主要的成员有IL-8、黑素瘤细胞生长刺激活性(GRO/MGSA)、血小板因子-4(PF-4)、血小板碱性蛋白、蛋白水解来源的产物CTAP-Ⅲ和β-thromboglobulin、炎症蛋白10(IP-10)、ENA-78;(2)C-C/β亚族,主要趋化单核细胞,这个亚族的成员包括巨噬细胞炎症蛋白1α(MIP-1α)、MIP-1β、RANTES、单核细胞趋化蛋白-1(MCP-1/MCAF)、MCP-2、MCP-3和I-309。
7.其它细胞因子 如表皮生长因子(EGF)、血小板衍生的生长因子(PDGF)、成纤维细胞生长因子(FGF)、肝细胞生长因子(HGF)、胰岛素样生长因子-I(IGF-1)、IGF-Ⅱ、白血病抑制因子(LIF)、神经生长因子(NGF)、抑瘤素M(OSM)、血小板衍生的内皮细胞生长因子(PDECGF)、转化生长因子-α(TGF-α)、血管内皮细胞生长因子(VEGF)等。
细胞因子_细胞因子 -受体
概念
细胞因子是由多种细胞产生的,具有广泛调节细胞功能作用的多肽分子,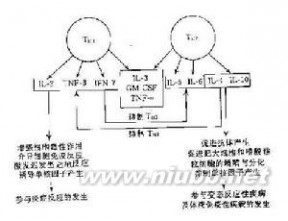 细胞因子生物学细胞因子不仅作用于免疫系统和造血系统,还广泛作用于神经、内分泌系统,对细胞间相互作用、细胞的增殖分化和效应功能有重要的调节作用。细胞因子发挥广泛多样的生物学功能是通过与靶细胞膜表面的受体相结合并将信号传递到细胞内部。因此,了解细胞因子受体的结构和功能对于深入研究细胞因子的生物学功能是必不可少的。随着对细胞因子受体的深入研究,发现了细胞因子受体不同亚单位中有共享链现象,这对阐明众多细胞因子生物学活性的相似性和差异性从受体水平上提供了依据。绝大多数细胞因子受体存在着可溶性形式,掌握可溶性细胞因子受体产生的规律及其生理和病理意义,必将扩展人们对细胞因子网络作用的认识。检测细胞因子及其受体的水平已成为基础和临床免疫学研究中的1个重要的方面。
细胞因子生物学细胞因子不仅作用于免疫系统和造血系统,还广泛作用于神经、内分泌系统,对细胞间相互作用、细胞的增殖分化和效应功能有重要的调节作用。细胞因子发挥广泛多样的生物学功能是通过与靶细胞膜表面的受体相结合并将信号传递到细胞内部。因此,了解细胞因子受体的结构和功能对于深入研究细胞因子的生物学功能是必不可少的。随着对细胞因子受体的深入研究,发现了细胞因子受体不同亚单位中有共享链现象,这对阐明众多细胞因子生物学活性的相似性和差异性从受体水平上提供了依据。绝大多数细胞因子受体存在着可溶性形式,掌握可溶性细胞因子受体产生的规律及其生理和病理意义,必将扩展人们对细胞因子网络作用的认识。检测细胞因子及其受体的水平已成为基础和临床免疫学研究中的1个重要的方面。结构和分类
根据细胞因子受体cDNA序列以及受体胞膜外区氨基酸序列的同源性和结构征,可将细胞因子受体主要分为4种类型:免疫球蛋白超家族(IGSF)、造血细胞因子受体超家族、神经生长因子受体超家族和趋化因子受体。此外,还有些细胞因子受体的结构尚未完全搞清,如IL-10R、IL-12R等;有的细胞因子受体结构虽已搞清,但尚未归类,如IL-2Rα链(CD25)。
(一)免疫球蛋白超家族
该家族成员胞膜外部分均具有1个或数个免疫球蛋白(Ig)样结构。已知,属于IGSF成员的细胞因子受体的IL-1RtI(CD121a)、IL-1RtⅡ(CD121b)、IL-6Rα链(CD126)、gp130(CDw130)、G-CSFR、M-CSFR(CD115)、SCFR(CD117)和PDGFR,并可分为几种不同的结构类型,不同IGSF结构类型的受体其信号转导途径也有差别。
(1)M-CSFR、SCFR和PDGFR:胞膜外区均含有五个Ig样结构域,其中靠近胞膜区为一个V样结构,其余四个为C2样结构。受体通常以二聚体形式与相应的同源二聚体配体结合。受体胞浆区本身含有蛋白酷氨酸激酶(proteintyrosinekinase,PTK)结构。
(2)IL-1RtI和IL-1RtⅡ:胞膜外区均含有三个C2样结构,受体胞浆区丝氨酸/苏氨酸磷酸化可能与受体介导的信号转导有关。
(3)IL-6Rα链、gp130以及G-CSFR:胞膜外区N端均含一个C2样区,在靠近胞膜侧各有一个红细胞生成素受体超家族结构域,此外在胞有胞膜外区还含有2-四个纤粘连素结构域。gp130胞浆区酷氨酸磷酸化与信号转导有关。这种结构类型的受体其相应配体IL-6、OSM、LIF和G-CSF在氨基酸序列和分子结构上也有很大的相似性。
(二)造血细胞因子受体超家族
造血细胞因子受体超家族(haemopoieticcytokinereceptorsuperfamily)又称细胞因子受体家族(cytokinereceptorfamily),可分为红细胞生成素受人本超家族(erythropoietinreceptorsuperfamily,ERS)和干扰素受体家族(interferonreceptorfamily)。
1.ERSERS所有成员胞膜外区与红细胞生成素(erythropoietin,EPO)受体胞膜外区体在氨基酸序列上有较高的同源性,分子结构上也有较大的相似性,故得名。
(1)ERS的成员:属于ERS的成员有EPOR、血小板生成素R、IL-2β链(CD122)、IL-2Rγ链、IL-3Rα链(CD123)、IL-3Rβ、IIL-4R(CDw124)、IL-5Rα链、IL-5βα链、IL-5Rβ链、IL-6Rα链(CD126)、gp130(CDw123)、IL-7R、IL-9R、IL-11R、IL-1240kDa亚单位、G-CSFR、GM-CSFRα链、GM=CSFRβ链、LIFR、CNTFR等,此外,某些激素如生长激素受体(GRGR)和促乳素受体(PRLR)亦属于ERS。
(2)ERS的结构特征:红细胞生成素受体超家族成员在胞膜外与配体结合部位有1个约含210氨基酸残基的牲性同源区域,主要特点①同源区靠近N端有四个高并能保守的半胱氨酸残基Cysl、Cys2、Cys3、Cys4和一个保守的钯氨酸,Cys1与Cys2之间、Cys3与Cys4之间形成2个二硫键。②同源区靠近细胞膜处,约在细胞膜外18~22氨基酸基处有1个色氨酸一丝氨酸-X-色氨酸-丝氨酸基序,所谓Trp-Ser-Xaa-Trp-Ser即WSXWS基序,其生物学功能尚不明了。IL-3α链、IL-3Rβ链、GM-CSFRβ链、LIFR办有2个ERS结构域,其中GM-CSFRβ链第1个ERS结构中有1个类似WSXWS基序,即为脯氨酸一丝氨酸-赖氨酸-色氨酸-丝氨酸(PSKWS)基序。1994年Hilton等合成WSXWS基序相应的寡核苷酸为探针,从成鼠肝cDNA文库中克隆小鼠IL-11受体α链cDNA获得成功。IL-6Rα链和gp130以及G-CSFrN端有1个IGSF结构。IL-7R靠近N端侧的部位只有Cys1和Cys3,与其它成员相比,缺乏Cys2和Cys4以及色氨酸残基。IL-1240kDa亚单位有ERS的同源结构,但为非膜结合的,而且与IL-12另一35kDa亚单位通过二硫键开成异源双体。GM-CSFrN端在ERS中可以看作由两个Ⅲ型纤维粘连素组成,每个Ⅲ型纤维粘连素结构域由7股反平行β折叠股形成1个桶状结构,2个桶状结构之间的槽是配体膜外保守区域有明显的进化同源性,这种同源性的程度与I
 细胞因子
细胞因子GSF成员间相似。EPoR似科与其它家族成员有更高的同源性,在进化上可能处于主导的地位。ERS的胞浆区长度不一,从5四个氨基酸残基到56八个氨基酸残基,除IL-2Rβ链与EPOR之间胞浆区有一定同源性外,其它成员在胞浆区未见明显的同源性。ERS成员胞浆区本身均不具备PTK结构,其信号传递的途径和机理也有所不同。IL-2Rβ链胞浆区的本双重性区与胞浆中酪氨酸激酶相关联,富含丝氨酸区与非激酶信赖途径有关。IL-2Rγ链胞浆区具有SH2结构,参与信号传递。细胞浆中的PTK和PKC可能参与IL-4R介导的信号传递。gp130胞浆区丝氨酸富含区以及酷氨酸磷酸化与gp130介导的信号转导有关。此外,酪氨酸磷酸化与IL-7R、GM-CSFRβ链、IL-3Rβ链、IL-5Rβ链介导的信号转导有关。
2.干扰素受体家族属于这一家族的成员有IFN-α/βR、IFN-γR和组织因子(TF)(为凝固白酶因子Ⅶ的细胞膜受体),其结构与红细胞生成素受体家族相似,但N端只含有2个保守性的Cys,2个Cys之间有七个氨基酸。近膜处也有2个保守的Cys,2个Cys之间间隔有20-二十二个氨基酸。IFN-α/βR由2个上述的结构域所组成。
(三)神经生长因子受体超家族
1.NGFR超家族的成员属于该家族成员除神经生长因子受体(nervegrowthfactorreceptorNGFR)外,不有TNF-RⅠ(CD120a)、TNF-RⅡ(CD120b)、CD40、CD27、T细胞cDNA-41BB编码产物、大鼠T细胞抗原OX40和人髓样细胞表面活化抗原Fas(CD95)。
2.NGFR超家族的结构特点NGFR超家族成员其胞膜外由3-六个约40个氨基酸组成的富含Cys区域,如NGFR、TNF-RⅠ、TNF-RⅡ有四个结构域,CD95有三个结构域,CD30有六个结构域。所有成员N端第1个区域中均含六个保守的Cys以及Tyr、Gly、Thr残基各1个,其它区域亦含4-六个Cys。TNF-RⅠ、CD95、CD40分子之间胞浆区约有40-50%同源性。
(四)趋化因子受体
1988年IL-8基因克隆成功以来,已形成了称之为趋化因子(chemokine)的1个家族。到目前为止,趋化因子家族的成员至少有十九个。部分趋化因子的受体已基本搞清,它们都性属于G蛋白偶联受体(GTP-bindingproteincoupledreceptor),由于此类受体有七个穿膜区,又称七个穿膜区受体超家族(sevenpredicatedtransmembranedomainreceptorsuperfamily,STRsuperfamily)。G蛋白偶联受体(或STR)包括的范围很广,除了趋化因子受体外,如某些氨基酸、乙酰胆硷、单胺受体,经典的趋化剂(C5a、fMLP、PAF)受体等都属于G蛋白偶联受体/STR。
1.趋化因子受体的种类和结构
(1)趋化因子受体的种类:已发现的趋化因子受体种类有IL-8RA、IL-8RB、MIP-1α/RANTEsR、NCP-1R和细胞趋化因子受体(redbloodcellchemokinereceptorRBCCKR)。有人将能与IL-8结合的IL-8RA、IL-8RB和RBCCKR(Duffy抗原)归为IL-8受体家族。
(2)趋化因子受体的结构:所有趋化因子受体都属于G蛋白偶联受体/STR,N端在胞膜外,C端位于胞浆内。七个穿膜区(transmembranedomain,TMD)为α螺旋,在TMDⅡ、Ⅳ、Ⅴ、Ⅵ和Ⅶ由α螺旋内保守的肺腑氨酸所扭结(kinked),胞膜外和胞浆内各有由亲水氨基酸所组成的3个一不,分别简称为e1-e3(e:extracellularconnectingloops)和il-i3(iintracellularconnectingloops)。e1和e2之间由2个保守的Cys形成1个二硫键,有些受体在胞外N端和e3之间也形成二硫键,如IL-8Ra30Cys与277ys形成二硫键。在STR超家族中,趋化因子受体以及经典的趋化剂受体具有以下特点:(1)其长度在STR超家族中最短,约为350氨基酸,其主要原因是N端、C端较短,i3环只含16-二十二个氨基酸;(2)在氨基酸水平上同源性大于20%;(3)i3富含碱性氨基酸,带正电;(4)N端仿酸,带负是电;(5)胞浆区含有多个丝氨酸和苏氨酸,可能是磷酸化位点;(6)mRNAs多表达于白细胞。
2.IL-8受体家族IL-8R家族是趋化因子受体中能与IL-8结合的不同受体的总称,包括IL-8RA、IL-8RB和RBCCKR。
(1)IL-8RA:IL-8RAcDNA1991年基因克隆成功,是Holmes等从中性粒细胞cDNA表达文库中分离得到,人IL-8RA基因定位于染色体2q35,与IL-8RB基因密切连锁和高度同源,可能是从同一祖先基因经复制而来。从cDNA推算出IL-8RA由350氨基酸组成,有五个N连接的糖基化位点。裸肽分子量为40kDa,糖基化后55~69kDa,在氨基酸水平上与IL-8RB的同源性为77%。IL-8RA只与配体IL-8(碱性,PI8.0-8.5)结合,这与IL-8RA的结构有关,IL-8RaN端酸性氨基酸是与IL-8结合的位置,N端Asp11和e3中Gly275和Arg280对于与配体结合至关重要,由于Cys30与Cys277之间形成二硫键,Asp11、Glu275和Arg280在空间置上十分接近,共同参与同配体的结合。IL-8RA基因表达的细胞种类较为广泛,如中性粒细胞、单核细胞、PGA活化的T细胞、单核细胞样细胞系、黑素瘤细胞、滑液成纤维细胞、HL60细胞和前髓样细胞系THP-1等。
(2)IL-8RB:IL-8RBcDNA是首先从HL60细胞中克隆成功,推断的氨基酸残基数为335,有1个潜在的N连接糖基化位点。IL-8RB可与CXC亚族中IL-8、GROα、GROβ、GROγ和NAP-2结合。人IL-8RB主要表达于髓样细胞,如中性粒细胞、HL60、THP-1和AML193细胞。
(3)RBCCKR:这种受体结合配体的特异性较宽,又称multi-specificreceptor,可结合CXC亚族中的IL-8、NAP-2、GROα和CC亚族中的MCP-1和RANTES。人RBCCKRcDNA1993年克隆成功,基因定位于1q21-q25,成熟受体分子由33八个氨基酸组成,分子量为39kDa,与IL-8RB和MIP-1α/RANTESR分别有27%和23%同源性。胞膜外区为6六个氨基酸,含有两个潜在的N连接糖基化点,酸性。C端胞浆区长二十四个氨基酸残基,RBC-CKR似乎不G蛋白调节,可能是1种G蛋白的非偶联受体。RBCCKR是人红细胞Duffy抗原(gpD),也是微小间日疟原虫(Plasmodiumvivax)受体。Duffy血型阴性个体尽管存在着该血型的原因,但不表达Duffy抗原/RBCCKR。RBCCKR作为1种清除受体(clearancereceptor)清除血液中趋化因子。这种受体与配体结合的亲和力Kd为5nM,正常血清中IL-8水平在pM水平。在成人呼吸窘迫综合征(ARDS)、脓毒症时,血清IL-8水平可升高至8nM,过高水平的IL-8结合到RBCCKR而得以清除。IL-8等趋化因子结合到红细胞上后即失去了对靶细胞作用。红细胞的这种清除作用的意义都还在于维持1个合适的趋化因子浓度,保证中性粒细胞等敏感地从血液中向趋化因子浓度高的炎症部位动。RBCCKR除表达在红细胞上外,还表达在肾脏、大脑,基因表在这还见于脾、肺和胸腺等。
3.受体的信号转导IL-8RA和IL-8RB中紧接第3个穿膜区(TMDⅢ)的第二个胞内环(i2)有一段高度保守的DRYLAIVHA序列,与受体信号的转导密切相关,其中DRY对于受体有效地偶联G蛋白是必要的,如用突变方法改变此序列,虽然不影响受体与配体的结合,但几乎完全丧失了配体刺激的生物学活性。IL-8R与配体结合后使与受体结合的异源三体G蛋白分解为α亚单位和βγ亚单位,α亚单位活化磷脂酶C(phospholipaseCPLC),导致胞浆内三磷酸肌醇(IP3)和二酰基甘油(DAG)增加,分别诱导胞浆内Ca库释放Ca2 和PKC的活化。此外,IL-8RA和IL-8RBC端丝氨酸和苏氨酸残基的磷酸化可能与信号的转导有关。
4.趋化因子受体与病毒最近发现某些感染人或灵长类病毒的开放读框产物与某些趋化因子受体有较高的同源性,这有可能与病毒的致病以及病毒所具有的某些生物学特性有关。
(1)人巨细胞病毒(humancytomegalovirusHCMV):是1种可感染人上皮细胞、髓样和淋巴样细胞的β疱疹病毒(βHerpesvirus)。HCMV三个开放读框US27、US28和UL33所推断的氨基酸序列在分子结构上均可模拟STR,其中US28产物与人MIP-1α/RANTEsR约有30%同源性,与该受体N端的同源性高达56%。US28产物可与趋化因子β亚族中MIP-1α、MIP-1β、MCP-1和RANTES相结合,但不能结合α亚族中的趋化因子。
(2)Saimiri疱疹病毒(HerpesvirussaimiriHVS):是1种感染灵长类动物嗜T细胞的γ疱疹病毒(γHerpesvirus)。HVS开放读框ECRF3产物与IL-8R有近30%的同源性,与IL-8RbN端的同源性为44%。ECRF3产物与IL-8、GROα和NAP-2均可发生一定程度的结合。
HCMV-US28和HVS-ECRF3探针不能与人基因组DNA杂交,提示疱疹病毒不仅从宿主体内获得了趋化因子受体基因拷贝,而且进行了修饰。类似的现象见于嗜人B淋巴细胞的γ疱疹病毒-EB病毒(EBV),EBV开放读框BCRF1是从宿主体内获得的IL-10基因,BCRF1产物又称为病毒IL-10(vIL-10),可模拟哺乳动物IL-10的抗炎症和抗增殖效应。
共享链
大多数细胞因子受体是由2个或2个以上的亚单位组成的异源二聚体或多聚体,通常包括1个特异性配体结合α链和1个参与信号的β链。α链构成低亲和力受体,β链一般单独不能与细胞因子结合,但参与高亲和力受体的形成和信号转导。应用配体竟争结合试验、功能相似性分析以及分子克隆技术发现在细胞因子受体中存在着不同细胞因子受体共享同1种链的现象。
(一)细胞因子受体共享链的种类
就在众多的细胞因子中,某些细胞因子的作用十分相似,如IL-3、IL-5、GM-CSF都作用于造血系统,促进造血干细胞或定向干细胞的增殖。IL-6、IL-11、LIF、OSM都能作用于肝细胞、巨核细胞、浆细胞瘤,发挥相似的生物学作用。IL-2、IL-4、IL-7、IL-9和IL-13均具有刺激T细胞或和B细胞增殖的作用。上述细胞因子功能的相似性已部分在受体水平得到解释,在很大程度上是由细胞因子受体共享链所决定的。目前已知,细胞因子共享链主要有gp310、GM-CSFRβ链和IL-2Rγ链。
1.gp130/LIFR为IL-6R、单抗MT18在骨髓瘤细胞系U266共沉淀中得到1种130kDa的糖蛋白,命名为gp130。1990年Hibi克隆成功,gp130,属于造血因子受体家族。IL-6、IL-11均能刺激IL-6信赖的小鼠浆细胞瘤系T1165的增殖,能在IL-3、GM-CSF的作用下缩短骨髓多能干细胞的Go期,增强IL-3依赖的人和小鼠的巨核细胞集落的形成,促进体内、体外的特异性抗体反应,诱导肝细胞急性期蛋白的产生。抗gp130能阴断IL-6、IL-112种细胞因子分别诱导的TF1细胞的增殖,而抗IL-5R只能阴断IL-6诱导的TF1的增殖,表明IL-6、IL-11受体共用1个信号转导链。OSM受体存在着低亲和力及高亲和力2种受体,低亲和力受体即gp130,gp130与LIFR构成高亲和力受体。与在IL-6R、IL-11R中不同,gp130在OSMR中只形成低亲和力受体且不能单独转导细胞因子信号。高亲和力的LIF受体由LIFR和gp130组成,OSM与LIF能竞争结合高亲和力LIF受体,但不竞争结合低亲和力的LIF受体。
(4)IL-11Rα链(小鼠)与IL-6Rα链和CNTFRα链氨基酸同源性分别为24%和22%。
2.KH97/AIC2B为IL-3R、IL-5R、GM-CSFR所共用。在造血方面,IL-3与GM-CSF均能促进未成熟细胞、混合细胞及粒细胞-巨噬细胞集落的形成,激活单核细胞,促进嗜酸性粒细胞集落形成。IL-5除了促进B细胞分化和分泌抗体外,也具有刺激嗜酸性粒细胞分化作用。用GM-CSFRβ链分别与IL-3、IL-5、GM-CSFRα链共转染的试验证明,这3种细胞因子高亲和力受体中的β链在小鼠和人分别为AIC2B和KH97,它们有56%的同源性。
3.IL-2受体γ链除IL-2R含有γ链外,IL-4R、IL-7R、IL-9R和IL-13R复合物中也共用IL-2Rγ链(γc)。这些受体的相应配体是一组主要作用于T细胞的生长因子。以IL-2γ链异常为主要特征的X联锁严重免疫缺陷综合症患者显示出T细胞发育异常,T细胞的缺乏或数量明显减少,提示IL-2γ链在T细胞的发育中起至关重要的作用。IL-4、IL-7均在T细胞的发育中起作用,它们共用一条信号转导链IL-2Rγ链来传递T细胞增殖的信号。在IL-2受体系统中,α链构成低亲和力受体,中亲和力受体由β、γ链组成,高亲和力受体由α、β、γ三条链组成,其中,γ链相当于其它细胞因子受体的β链,参参与信号传递,而αβ链则相当于α链,主要发挥识别和结合配体的作用。
(二)共享链与细胞因子受体信号转导
细胞因子信号转导首先需要配体与受体结合并诱导受体二聚体(或三聚体)的形成,使二聚体(或三聚体)胞浆部分的相互作用,由此引起不同途径的信号转导。在IL-2R系统中,受β、γ链的二聚作用对于信号的转导是必须的,缺乏β链胞浆区的IL-2R不能转导IL-2刺激所发生的信号。大多数的细胞因子对细胞的刺激及信号转导与酪氨酸激酶的活化及细胞内蛋白的酪氨酸磷酸化有关,细胞因子与受体结合可以引起受体成分的酪氨酸磷酸化。ERS胞浆区近膜端的60个氨基酸残基是高度保守的,这段同源序列对IL-6、G-CSF、EPO、IL-7的信号转导起着关键作用,提示这些受体可能利用相似的胞膜内信号转导机制。
1.gp130介导信号转导在IL-6R、IL-11R、OSMR、LIFR、CNTFR的信号转导共用链gp130中,其胞浆区约27七个氨基酸残基中包含丝氨酸富含区、核苷酸结合区及四个GTP结合模式区。其中的丝氨酸富含区也存在于G-CSFR、IL-2Rβ、IL-4R和EPOR,其它的ERS成员有着明显的同源性。其中1个片段在所有ERS成员中都是保守的,另1个片段存在于G-CSFR、EPOR、KH97中。这2个短的片段中,无论哪个发生突变都将使gp130不能发生酪氨酸磷酸化,丧失信号转导的能。LIFR/gp130异源双体也与酪氨酸磷gp130不能发生要酪氨酸磷酸化,丧失信号转导的功能。LIFR/gp130异源双体也与酪氨酸磷酸化有关。虽然大多数造血因子受体家族成员均不具有酪氨酸激酶结构域,但它们与酪氨酸激酶型生长因子受体相似,生长因子引起与之相关的受体酪氨酸激酶二聚体的形成和激活,而造血因子可能是诱导其受体的二聚体形成并导致相关酪氨酸激酶的活化。已发现在IL-6、IL-11刺激的TF1细胞中检测出分子量97/95kDa的蛋白发生了酪氨酸磷酸化,抗gp130的信号转导中很重要。在不同的细胞系3T3-L1、B细胞杂交瘤、髓样白血病系中发现有不同分子量蛋白的要酪氨酸磷酸化,提示在不同的细胞系
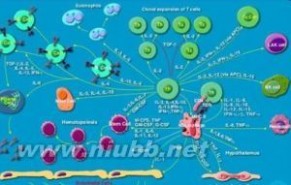 细胞因子
细胞因子中存在细胞特异的酪氨酸激酶及各自特异的底物,这有可能是共用gp130的IL-6、IL-11、LIF、CNTF、OSM在不同细胞中生物学作用差异的原因一。JAK2是1种非受体型的酪氨酸激酶,可以被EPO、IL-3、G-CSF、IL-6等多种细胞因子刺激所激活,JAK2可能是这些不同的细胞因子受体信号转导途径中的1个共同因素,这种与受体相联的JAK2激酶可能因受体结构的不同而催化不同的底物,从而导致了JAK2介导了许多不同的生和的学功能。此外,gp130在IL-6、IL-11、CNTF、LIF的刺激后也发生了自身的酪氨酸磷酸抡。
2.KH97/AIC2B介导信号传导在IL-3、IL-5、GM-CSF的信号转导链KH97/AIC2B的胞浆区内也存在着2个产生不同信号所必需的区域:1个是Glu517上游近膜端的约60个氨基酸的区域,它是诱导c-myc和pim-1所必需的;另1个区域是Leu623至Ser763约140个氨基酸的胞浆区域,是Ras、Raf、MAP(丝裂原激活的蛋白激酶)的激活以c-fos、c-jun的诱导所必需的。hGM-CSFRα、β链无任何已知酶的催化区,共转染hGM-CSFα、β链的Ba/F3细胞的地冽同C-Myc、pim-1水平的延长增加相关联的。在小鼠淋巴细胞系转染GM-CSFα、β链后可以引韦胞内数种蛋白的酪氨酸磷酸化并引起增殖反应,α、β链共转染小鼠NIH3T3细胞表达GM-CSFR高亲和力受体,可引起表达的β链胞浆区和另外1个有包浆内40-45kDa蛋白的酪氨酸快速磷酸化。
可溶性
在自然关态下,细胞因子受体(cytokinereceptorCK-R)主要以膜结合细胞因子受体(membrane-boundcytokinereceptorMCK-R)和存在于血清等体液中可溶性细胞因子受体(solublecytokinereceptorsCK-R)2种形式存在。细胞因子复杂的生物学活性主要是通过基与相应的mCK-R结合后所介导的,而sCK-R却具有独特的生物学意义。sCK-R水平变化与某些疾病的关系日益受到学者们的重视。部分重组sCK-R(rsCK-R)基因工程产品已进入临床验证,关于sCK-R的产生机理,结构特点及基免疫学功能等方面的基础研究也取得了长足的进展。
(一)sCK-R的产生机理及作用特点
人T细胞白血病病毒I型(HTLV-I)感染的HUT102B2、MT-2等细胞,髓样白血病细胞(HL-60,KG1)及某些人B细胞系(Raji)除了表达多种mCK-R外还可以通过不同方式产生sCK-R,如HUT102B2细胞培养丰清中也可检出高水平sCK-2R和sIL-6R。人PMC体外经PHA刺激培养后也产生大量sCK-2R和sCK-6R。
1.sCK-R的产生大多sCK-R主要来自膜受体的脱落,因此将膜受体阳笥细胞溶解是获得大量sCK-R的1种方法。大多数sCK-R氨基酸序列与mCK-R胞膜外区同源,只缺少跨膜区及胞浆区,但仍可与相应配体发生特异性结合所应。除了膜受体的裂解、脱落产生sCK-R形式外,另1种产生sCK-R的机理是通过受体mRNA不同剪接,产生分泌型mRNA,通过翻译后直接分泌到细胞外,已经证实,细胞内可含有同1种CK-R不同形式的cDNA。sIL-4R、sIL-5Rα链、sIL-6Rα链、sIL-7R以及sG-CSFR等可以这种形式产生。
2.sCK-R的生物学作用多数sCK-R与相应细胞因子结合的亲和力较与mCK-R为低,可能与sCK-R为单链结构或缺少某些结构区域有关。也有的sCK-R如sIL-4R同天然mIL-4R与相应配体结合的亲和力是相同的,即使低剂量sIL-4R也可特异地抑制IL-4诱导的细胞增殖反应。sCK-R以其多种方式发挥其独特的免疫学功能。
(1)做为细胞因子转运蛋白,将细胞因子运至机体有关产啊位,造成局部细胞因子高浓度区以充分发挥细胞因子的生物学效应。
(2)是膜受体正常代谢途径,有利于处于活化状态细胞恢复至正常水平。
(3)竟争性地结合mCK-R相应配体,抑制mCK-R所介导的生物学作用。
(二)sCK-R与临床
1.检测sCK-R水平在临床中的应用检测某些sCK-R水平辅助临床对某些疾病的早期诊断,了解病程的发展与转归,并可对患者免疫功能状态及预后进行评估,对临床治疗也有一定指导意义。
(1)sIL-2R的检测:来国内外学者对sIL-2R进行了大量研究,发现其在血清及其他体不认中水平的变化与临床多种疾病如器官移植排异反应、病毒性感染、恶性肿瘤、创伤及自身免疫性疾病等的病情、病程密切相关。
(2)其他sCK-R的检测:最近在尿中发现1种50kDasIL-6R分子称为IL-6R-SUP,可以促进小剂量IL-6诱导的小鼠浆细胞瘤T1165的生长。多发性骨髓瘤病人血浆中sIL-6R水平明显升高。风湿性疾病患者血清sTNFR水平异常增设,关工了腔滑液中亦可检出高水平sTNF-R,且活动期水平明显高于非活动期。政党妇女尿中仅可检测到TNF-RⅡ类型的sTNFR,而妊娠妇女尿中TNF-rI和TNF-RⅡ2种类型sTNFR均可被检出,随胎龄增加sTNFR水平逐渐升高,分娩后随之降低,这有可能是使胎儿免受TNF作用的1种防护机制。肝脏感染性腹水及癌性腹水中亦可检出高水平sTNFR。此外还发现,sTNFR水平的增设与患者肾功能的减退密切相关。
2.sCK-R在临床应用前景大多数sCK-R与细胞因子结合后阴断细胞因子与膜受体结合,从而抑制细胞因子的生物学活性,应用sCK-R减轻或防止炎症性细胞因子造成的病理损害提供了新的治疗途径。动物实验结果表明,局部注射sIL-1R可抑制IL-1介导的炎症反应。sIL-1R可降低小鼠同种异体心脏移植的排异反应以及大鼠实验性关节炎和过敏性大脑炎。在体外sIL-1R可明显抑制急性髓样白血病病人骨髓细胞的增殖。最近,应用IL-1R基因工程产品开始对治疗关节炎、糖尿病以及防治器官移植排斥等进入临床验证。动物体内注射sIL-4R可延长同种异人本移植物的存活,抑制GVHR,降低I型超敏反应。应用sTNFR可减轻TNF在自身免疫性疾病中所介导的病理损害,并可减轻败血症休克。
细胞因子_细胞因子 -作用特点
众多的细胞因子有以下共同的作用特点。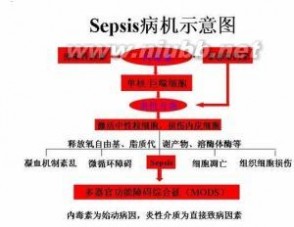 细胞因子连锁
细胞因子连锁(1)绝大多数细胞因子为分子量小于25kDa的糖蛋白,分子量低者如IL-8仅8kDa。多数细胞因子以单体形式存在,少数细胞因子如IL-5、IL-12、M-CSF和TGF-β等以双体形式发挥生物学作用。大多数编码细胞因子的基因为单拷贝基因(IFN-α除外),并由4-五个外显子和3-四个内含子组成。
(2)主要与调节机体的免疫应答、造血功能和炎症反应有关。
(3)通常以旁分泌(paracrine)或自分泌(autocrine)形式作用于附近细胞或细胞因子产生细胞本身。在生理状态下,绝大多数细胞因子只有产生的局部起作用。
(4)高效能作用,一般在pM(10-12M)水平即有明显的生物学作用。
(5)存在于细胞表面的相应高亲和性受体数量不多,在10-10000/每个细胞。细胞因子受体的研究进展相当迅速,根据细胞因子受体基因DNA序列以及受体胞膜外区氨基酸序列、同源性和结构,可分为4个类型:免疫球蛋白超家族、造血因子受体超家族、神经生长因子受体超家族和趋化因子受体。
(6)多种细胞产生,1种IL可由许多种不同的细胞在不同条件下产生,如IL-1除单核细胞、巨噬细胞或巨噬细胞系产生外,B细胞、NK细胞、成纤维细胞、内皮细胞、表皮细胞等在某些条件下均可合成和分泌IL-1。
(7)多重的调节作用(multipleregulatoryaction),细胞因子不同的调节作用与其本身浓度、作用靶细胞的类型以及同时存在的其它细胞因子种类有关。有时动物种属不一,相同的细胞因子的生物学作用可有较大的差异,如人IL-5主要作用于嗜酸性粒细胞,而鼠IL-5还可作用于B细胞。
(8)重叠的免疫调节作用(overlappingregulatoryaction),如IL-2、IL-4、IL-9和IL-12都能维持和促进T淋巴细胞的增殖。
(9)以网络形式发挥作用,细胞因子的网络作用主要是通过以下3种方式:(1)1种细胞因子诱导或抑制另1种细胞因子的产生,如IL-1和TGF-β分别促进或抑制T细胞IL-2的产生;(2)调节同1种细胞因子受体的表达,如高剂量IL-2可诱导NK细胞表达高亲和力IL-2受体;(3)诱导或抑制其它细胞因子受体的表达,如TGF-β可降低T细胞IL-2受体的数量,而IL-6和IFN-γ可促进T细胞IL-2受体的表达。
(10)与激素、神经肽、神经递质共同组成了细胞间信号分子系统。
细胞因子_细胞因子 -原理
为了维持机体的生理平衡,抵抗病原微生物的侵袭,防止肿瘤发生,机体的许多细胞,特别是免疫细胞合成和分泌许多种微量的多肽类因子。它们在细胞之间传递信息,调节细胞的生理过程,提高机体的免疫力,在异常情况下也有可能引起发烧、炎症、休克等病理过程。这样1大类因子已发现的有上百种,统称为细胞因子,包括淋巴细胞产生的淋巴因子、单核细胞产生的单核因子、各种生长因子等。许多细胞因子是根据它们的功能命名的,如白细胞介素(IL)、干扰素(IFN)、集落刺激因子(CSF)、肿瘤坏死因子(TNF)、红细胞生成素(EPO)等。
细胞因子_细胞因子 -作用
细胞因子通过结合细胞表面相应的细胞因子受体而发挥生物学作用。细胞因子与其受体集合后启动复杂的细胞内分子间的相互作用,最终引起细胞基因转录的变化。
参与免疫应答与免疫调节,调节固有免疫和适应性免疫应答;刺激造血功能;刺激细胞活化、增殖和分化;诱导或抑制细胞毒作用,诱导其凋亡。
细胞因子
细胞因子的作用方式:1.自分泌作用。2.旁分泌作用。3.内分泌作用。
细胞因子的作用特点:多效性、重叠性、协同性、拮抗性和双重性。
细胞因子研究具有非常重要的理论和实用意义,它有助于阐明分子水平的免疫调节机理,有助于疾病的预防、诊断和治疗,特别是利用基因工程技术生产的重组细胞因子已用于治疗肿瘤、感染、炎症、造血功能障碍等,并收到良好疗效,具有非常广阔的应用前景。
细胞因子_细胞因子 -结构
从分子结构来看,细胞因子都是小分子的多肽,多数由100个左右氨基酸组成。细胞因子都是通过与靶细胞表面的细胞因子受体特异结合后才能发挥其生物学效应,这些效应包括促进靶细胞的增殖和分化,增强抗感染和杀肿瘤细胞效应,促进或抑制其他细胞因子的合成,促进炎症过程,影响细胞代谢等。细胞因子的这些作用具有网络性的特点,即每种细胞因子可作用于多种细胞;每种细胞可受多种细胞因子的调节;不同细胞因子之间具有相互协同或相互制约的作用,由此构成了复杂的细胞因子免疫调节网络。人们对这一网络的认识尚远未清晰明了。
细胞因子_细胞因子 -研发历史
最近几年,基因重组的细胞因子作为1种新型的生物应答调节剂在临床应用上取得了令人瞩目的成就。例如,最早用于临床的干扰素α在治疗白血病和病毒感染中收到显著疗效。中国的干扰素a1在1991年通过新药审评,已得到较为广泛的应用。在国际上已批准生产的细胞因子药物还包括EPO、干扰素γ、GM-CSF、G-CSF、IL-2等。由于细胞因子为人体自身成分,通过调节机体生理过程和提高免疫力来治疗疾病,在低剂量就可以发挥作用,因而疗效显著,副作用小是1种全新的生物疗法,将会很快获得突飞猛进的发展。
细胞因子的研究渊源始于50年代的干扰素研究和60年代的集落刺激因子研究,由于基因工程技术的迅速发展,使细胞因子研究发生了突破性的进展。在80--90年代相继克隆出1大批细胞因子,细胞因子的化学本质是多肽,从信息传递的角度,细胞因子是生物体内1类重要的第一信使分子,是细胞内基因表达的产物。
在自然状态下,细胞因子受体(cytokinereceptor,CK-R)主要以膜结合细胞因子受体(membrane-boundcytokinereceptor,mCK-R)和存在于血清等体液中可溶性细胞因子受体(solublecytokinereceptor,sCK-R)2种形式存在。细胞因子复杂的生物学活性主要是通过其与相应的mCK-R结合后所介导的,而sCK-R却具有独特的生物学意义。sCK-R水平变化与某些疾病的关系日益受到学者们的重视。部分重组sCK-R(rsCK-R)基因工程产品已进入临床验证,关于sCK-R的产生机理,结构特点及其免疫学功能等方面的基础研究也取得了长足的进展。
研究表明,体内的各种细胞因子之间并不是孤立存在的,而是有着复杂的相互作用,它们之间通过合成和分泌的相互调节,受体表达的相互调节、生物学效应的相互影响等组成1个复杂的细胞因子互作网络。
二 : 白蛋白主要生理作用 偏低危害及偏高原因
白蛋白主要生理作用:
[www.61k.com)维持血浆胶体渗透压的恒定:白蛋白是血浆中含量最多、分子最小、溶解度大、功能较多的一种蛋白质。血浆胶体渗透压的维持主要依靠血浆中的白蛋白,胶体渗透压是使静脉端组织间液重返回血管内的主要动力。当血浆白蛋白因病理条件引起下降时,血浆的胶体渗透压也随之下降,可导致血液中的水份过多进入组织液而出现水肿。
血浆白蛋白的运输功能:血浆白蛋白能与体内许多难溶性的小分子有机物和无机离子可逆地结合形成易溶性的复合物,成为这些物质在血液循环中的运输形式。由此可见白蛋白属于非专一性的运输蛋白,在生理上具有重要性,与人体的健康密切相关。
浆白蛋白的其它生理作用:血浆中白蛋白的含量远比球蛋白多,亲水作用又比球蛋白大,这使血浆中的白蛋白对球蛋白起到一种胶体保护的稳定作用。当肝脏功能障碍引起白蛋白合成不足时,可使血浆球蛋白失去胶体保护作用,稳定性下降。血浆球蛋白的稳定性下降,将严重影响这些物质在体内的代谢、利用,引起相应的症状。此外,白蛋白还是人体内一种重要的营养物质。白蛋白在血浆中也不断地进行着代谢更新,血浆白蛋白分解产生的氨基酸,可用于合成组织蛋白,氧化分解以供应能量或转变成其它含氮物质。
白蛋白的功效:具有活性的激素或药物当与白蛋白结合时,可以不表现其活性,而视为其储存形式,由于这种结合的可逆性和处于动态平衡,因此在调节这些激素和药物的代谢上,具有重要意义。
白蛋白是具有黏性、胶质性的物质,在人体内遇到重金属离子时,会自动与重金属离子结合,由排泄系统排出体外,起到解毒的作用。因此,食用含白蛋白丰富的食物,可避免重金属离子的吸收而中毒。白蛋白对胃壁还有保护功效。
人血白蛋白是血液制品的一种,俗称“生命制品”、“救命药”。它是从健康人的血液中提炼加工而成,直接静脉注射到病人体内,其主要功能是增强人的免疫力和抵抗力。市场价300到400元一瓶。临床上主要用于失血创伤和烧伤等引起的休克、脑水肿,以及肝硬化、肾病引起的水肿或腹水等危重病症的治疗,以及低蛋白血症病人。
具有活性的激素或药物当与白蛋白结合时,可以不表现其活性,而视为其储存形式,由于这种结合的可逆性和处于动态平衡,因此在调节这些激素和药物的代谢上,具有重要意义。
白蛋白偏低的原因有很多种,比如胶体渗透压紊乱出现水肿氨基酸、激素、维生素等器官供应减少,分解代谢产物不能及时排泄出去,都可以导致白蛋白偏低。在临床诊疗中,白蛋白偏低的危害主要表现在两个方面:
1、当肝功能受损时,白蛋白产生减少,其降低程度与肝炎的严重程度是相平行的。慢性和重型肝炎及肝硬化患者血清白蛋白浓度降低。
2、白蛋白的作用之一是营养细胞和维持血管内的渗透压。当白蛋白减少时,血管内渗透压降低,患者可出现腹水。
一般情况下,肝脏疾病患者在肝功能检查中白蛋白偏低的现象比较常见,因为肝脏疾病患者往往存在肝损伤,导致白蛋白产生减少。因此,肝脏疾病患者要定期检查白蛋白,根据白蛋白降低程度了解病情轻重,及时进行治疗,防止肝腹水、肝硬化等肝脏疾病的发生。
白蛋白高的原因:
一:白蛋白高主要见于血液浓缩而导致相对性的增高,比如:严重的脱水和休克,大量出血、严重的烧伤、肾脏疾病等。
二:此外白蛋白高也可受到饮食中蛋白质摄入量影响,在一定程度上可以作为个体营养状态的评价指标。有时血液浓缩时可导致蛋白浓度相对增高,腹泻、呕吐、高热时急剧失水也可导致血清中白蛋白浓度增高,从而引起白蛋白高。
三 : 生活小贴士白细胞高的原因及危害 不容忽视的健康大问题
白细胞高的原因及危害
白细胞增多是指血液中白细胞比例增高的现象一般见于急性感染白细胞是血液中一类细胞的总称主要包括单核细胞淋巴细胞中性粒细胞白细胞的正常值是-x-/ml高于这个范围称为白细胞的增多常见于急性感染慢性感染等各种细菌或病毒感染性疾病有炎症的时候血液尿液里面的白细胞就会增加还有一些血液病的白细胞也会增加。(www.61k.com)此外医生常常要根据血常规中的白细胞状况进行分析判断患者是否存在感染一般来讲感染会引起白细胞增高但某些白细胞增高不一定就是存在感染许多生理因素可以引起白细胞总数增加比如:剧烈运动;体力劳动;在冬季长时间暴露于冷空气之后;饱餐淋浴后也常有白细胞轻微增高生理性白细胞增高还见于情绪紧张饥饿时低血糖等但生理性白细胞增多是暂时的去除影响因素则很快恢复产生机制可能是与各种生理因素刺激时体内儿茶酚胺分泌增多导致边缘白细胞进入循环所致许多药物可以引起白细胞总数增加比如:某些抗生素如红霉素头孢赛曲等;还有儿茶酚胺类药如肾上腺素去甲肾上腺素间羟胺多巴胺等;另外肾上腺皮质激素氢化可的松地塞米松促肾上腺皮质激素等均可引起白细胞总数增多抗精神病药碳酸锂也可以引起白细胞数量增多总之血常规中白细胞增高不一定是感染很可能是由于上述生理因素或药物所致血液病的白细胞也会增加。
白细胞高的原因及危害
白细胞数值:成年人外周血液中正常的白细胞数值是4~10×109/l,如果少于4×109/l为白细胞减少,超过10×109/l及为白细胞增多。引起白细胞与粒细胞增多的原因有:1生理性增多:见于新生儿、妊娠、剧烈运动、寒冷刺激等。2病理性增多:急性感染、严重创伤、急性出血、急性中毒、白血病、恶性肿瘤等。指导意见:白细胞增多本是一种防御性反应,在防御感染中起重要作用。但如果白细胞由于某种原因而异常增生,则会引发白血病,对机体造成发热、继发感染、贫血、出血、组织坏死、代谢及功能障碍等,甚至危及生命。 血液中的白细胞(wbc)是人体血液中具有防御和免疫功能的重要细胞,它来源于骨髓中的原始和幼稚血细胞,在骨髓里发育为成熟细胞后释放到血流中。按国际标准制,要求用每升中的数量来表示,为(4~9)×109/升。指导意见:白细胞的生理性升高常在进餐后、激烈运动后或受精神打击后,以及女性的月经期、妊娠后期等,但通常不会高于14×109/升。白细胞的病理性升高的原因很多,主要是体内有组织损伤或有炎症。 生理性血白细胞升高,没有危害的;还有病理性血白细胞增高,是身体或组织病变的一种临床症状。像某些细菌性感染所引发的疾病,特别是化脓性球菌引起的局部炎症和全身性感染,还有某些病毒性感染所导致的疾病,都会引起血白细胞会非正常升高,严重的组织损伤或坏死、过敏反应、中毒反应、肿瘤及血液病,慢性粒细胞白血病,急性粒细胞白血病等,也都会引起血白细胞会非正常升高,白细胞偏高说明机体有炎症发生。白细胞有多种,请问你是哪种白细胞偏多?是嗜中性粒细胞,还是嗜酸性粒细胞,还是淋巴细胞?嗜中性粒细胞多说明有炎症反应,就是说身体内可能有损伤比如出血,血栓或肿瘤什么的。嗜酸性粒细胞多说明身体内有寄生虫或细菌感染。淋巴细胞多说明有病毒或细菌感染,或感染后期机体内产生免疫力正在产生抗体。
白细胞高的原因及危害
医生常常要根据血常规中的白细胞状况进行分析,判断患者是否存在感染。一般来讲感染会引起白细胞增高。但某些白细胞增高不一定就是存在感染。许多生理因素可以引起白细胞总数增加。比如:剧烈运动;体力劳动;在冬季长时间暴露于冷空气之后;饱餐、淋浴后也常有白细胞轻微增高。生理性白细胞增高还见于情绪紧张,饥饿时低血糖等。但生理性白细胞增多是暂时的,去除影响因素则很快恢复。产生机制可能是与各种生理因素刺激时,体内儿茶酚胺分泌增多,导致边缘白细胞进入循环所致。许多药物可以引起白细胞总数增加。比如:某些抗生素如红霉素、头孢赛曲等;还有儿茶酚胺类药如肾上腺素、去甲肾上腺素、间羟胺、多巴胺等;另外肾上腺皮质激素氢化可的松、地塞米松、促肾上腺皮质激素等均可引起白细胞总数增多。抗精神病药碳酸锂也可以引起白细胞数量增多。 总之,血常规中白细胞增高不一定是感染,很可能是由于上述生理因素或药物所致。
本文标题:白细胞低的原因及危害-细胞因子:细胞因子-概述,细胞因子-重组要求及状态61阅读| 精彩专题| 最新文章| 热门文章| 苏ICP备13036349号-1